
肺癌是最常见的肺部原发性恶性肿瘤,死亡率居所有癌种首位,已成为我国第一大癌症。非小细胞肺癌(NSCLC)为肺癌的常见分型,约占所有肺癌病例的80%-85%[1],目前发现其发生发展机制与某些驱动基因突变相关,EGFR突变是NSCLC最常见的驱动基因之一,特别是在亚洲人群中,此类突变的发生率较高。
2024年8月强生宣布其EGFR/MET双抗amivantamab-vmjw(埃万妥单抗)与第三代EGFR抑制剂lazertinib(拉泽替尼)联用方案经美国FDA批准上市。用于一线治疗晚期EGFR突变的非小细胞肺癌患者,是目前唯一一个与奥希替尼相比,显示出更优的无进展生存期(PFS)的无化疗方案。
EGFR 是表皮生长因子受体家族成员之一,是一种跨膜糖蛋白, 具有配体诱导酪氨酸蛋白激酶活性, 广泛分布于除血管组织外的上皮细胞膜上。EGFR 与肿瘤细胞增殖、 血管生成、 肿瘤侵袭、 转移及细胞凋亡抑制有关。 研究表明, EGFR 在多种实体瘤中高表达或异常表达, 如胶质细胞瘤、 肺癌、 前列腺癌、 胰腺癌等肿瘤组织中都有 EGFR 过度表达。目前已有多种EGFR靶向治疗药物获批上市,包括单抗、小分子及双抗等药物。
EGFR单抗药物
靶向细胞表面的特异性抗原EGFR的单克隆抗体,可以通过多种机制抑制肿瘤生长,其主要抗肿瘤机制如下:
- 竞争性阻断配体与EGFR结合;
- 在空间上阻碍EGFR与其他家族成员的结合;
- 促进EGFR内化和降解;
- 激活补体依赖性细胞毒性(CDC)通路;
- 导致细胞周期阻滞于G1;
- 抑制促血管生成因子的表达;
- 促进细胞凋亡;
- 自然杀伤(NK)细胞或巨噬细胞介导的抗体依赖性细胞毒性(ADCC);
- 增加H2AX水平,抑制DNA-pk。
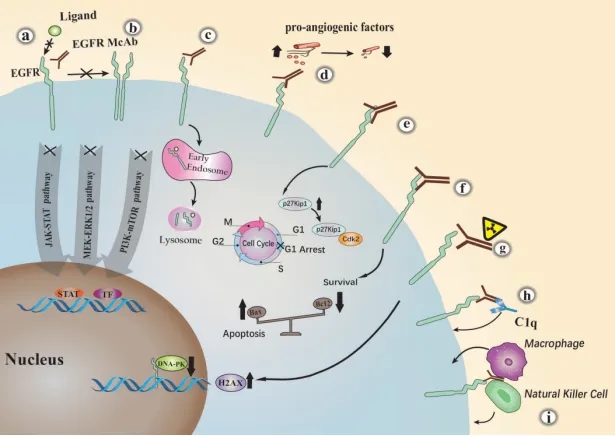
EGFR抗体主要抗肿瘤机制[2]
EGFR-TKI药物
针对EGFR突变的酪氨酸激酶抑制剂(TKIs)已经成为EGFR突变肺癌患者的一线治疗选择,EGFR-TKI药物近年来一直在更新换代,目前上市的药物已经发展到第三代。然而,尽管初始治疗反应良好,但几乎所有接受EGFR TKI治疗的患者最终都会不可避免的发展出获得性耐药,导致疾病进展生存受限。
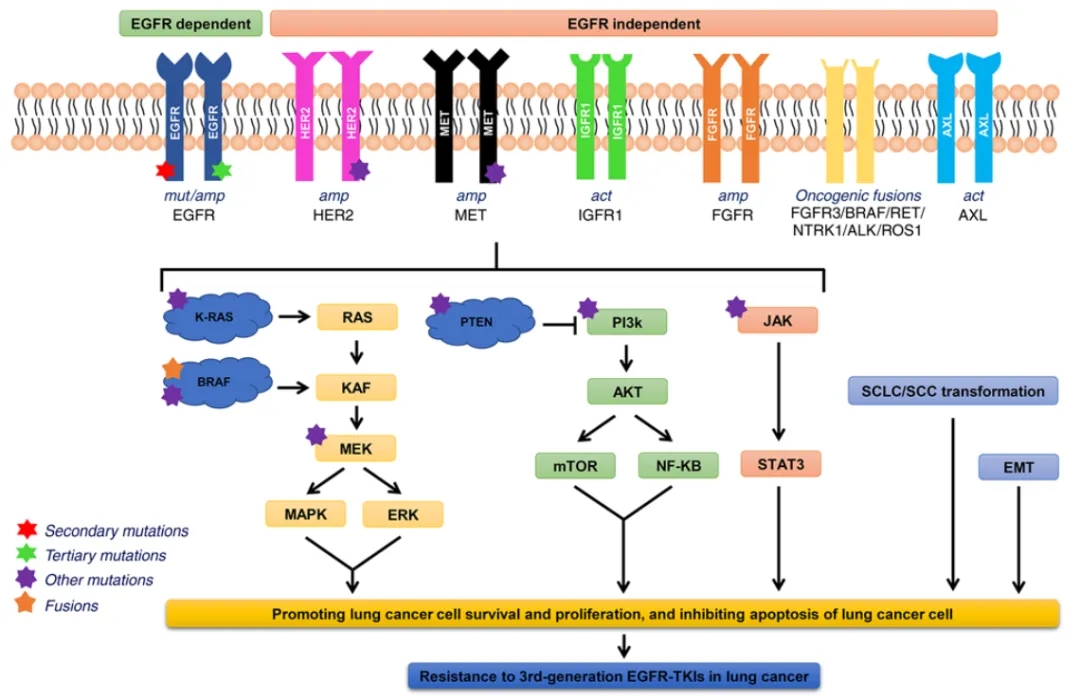
三代EGFR-TKI的获得性耐药机制[3]
进一步研究发现,合并原发间质-上皮细胞转化因子(MET)异常是导致EGFR突变患者对EGFR-TKI治疗无反应或反应时间较短的重要原因[4]。MET(或c-MET)基因扩增一直被认为是第一代或第三代EGFR-TKls的重要耐药机制。联合靶向治疗是耐药患者的一种很有前景的策略,多种新型联合治疗的临床研究正在进行中。
MET信号通路
MET(c-MET,cellular-mesenchymal epithelial transition factor))是细胞间质上皮转换因子,受体酪氨酸激酶家族成员,是调节上皮癌起始、进展和恶性肿瘤的致瘤性决定因素。MET信号通路激活可介导的细胞生长、存活、侵袭性、化疗耐药和致瘤性等。MET 在一般组织中呈低表达或不表达,但在多种类型的实体瘤中如肺癌、胃癌、肝癌、乳腺癌、皮肤癌、大肠癌等存在异常表达或突变,在肿瘤的发生发展中发挥了重要的作用。
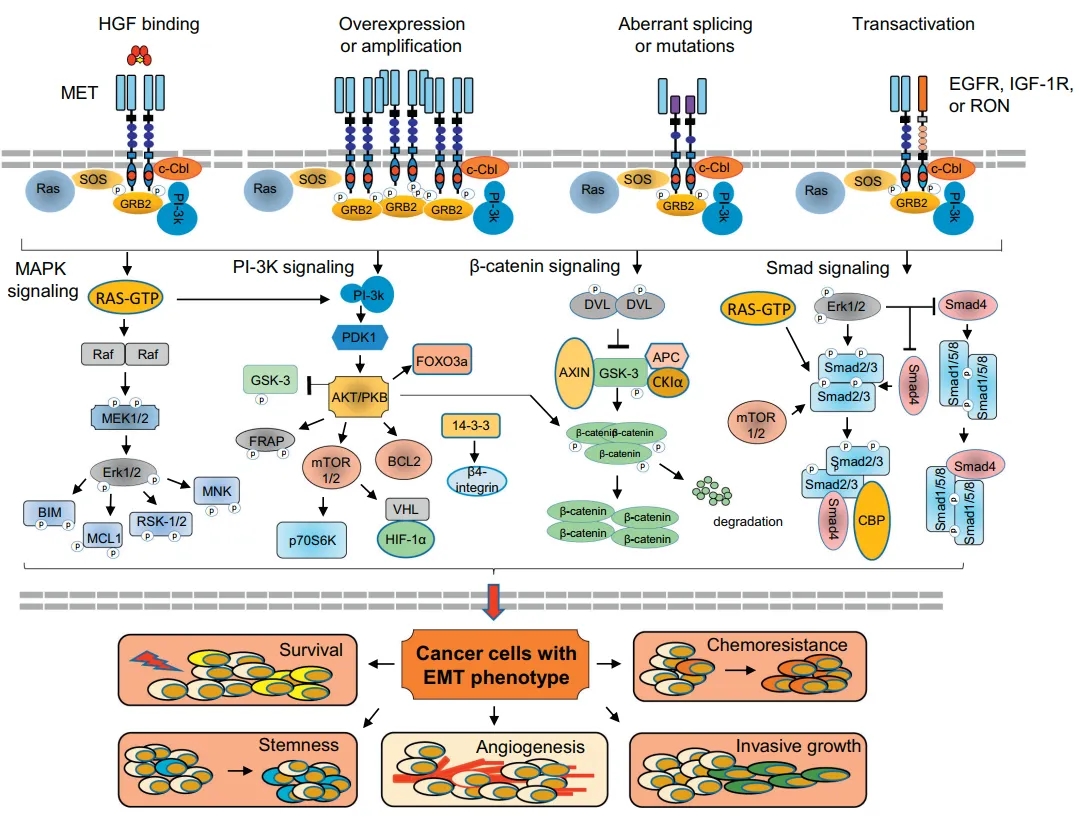
MET的激活机制、信号通路和生物学后果[5]
研究表明,MET被激活后,诱导细胞内酪氨酸残基的同二聚化和磷酸化,从而激活下游RAS/ERK/MAPK和PI3K-AKT信号通路。这些通路可以驱动细胞存活、增殖、运动、迁移、侵袭、血管生成和上皮-间质转化(EMT),在EGFR抑制剂存在下提供了一条旁路通路,导致耐药性增加[6,7]。
靶向EGFR/MET双特异性抗体
靶向双肿瘤相关抗原的双特异性抗体(BsAb)具有调用两种信号通路之间的协同效应、增加靶组织特异性和降低全身毒性的优势。通过结合抗体介导的特异性靶向和细胞毒性有效载荷的有效杀伤,抗体-药物偶联物(ADC),特别是双特异性ADC (BsADC),已成为强大的治疗策略。
EGFR和MET是在多种肿瘤中共同表达的致癌蛋白。此外,MET扩增与非小细胞肺癌(NSCLC)患者EGFR酪氨酸激酶抑制剂的耐药有关。这些结果表明EGFR/MET双特异性抗体可以成为EGFR和MET共表达肿瘤的有效治疗选择,并克服MET驱动的EGFR-TKI耐药,从而改善患者预后。
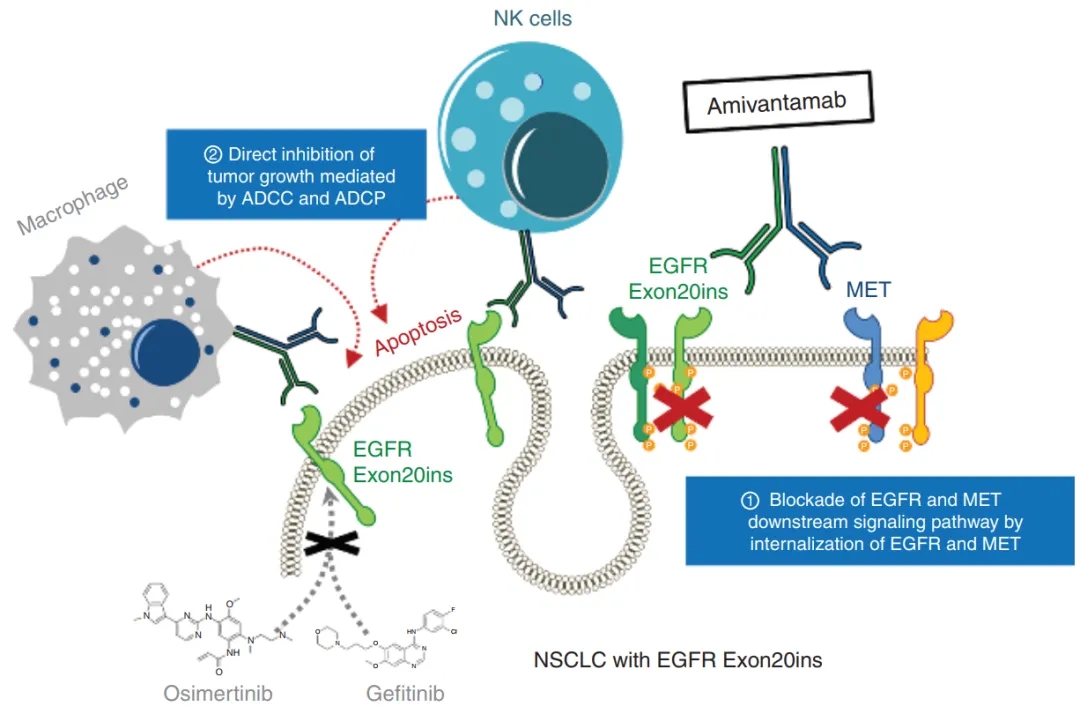
EGFR/MET双特异性抗体抗肿瘤机制[8]
百奥动物开发了B-hEGFR/hMET、B-hEGFR、B-Tg(hEGFR) MC38等一系列人源化小鼠及细胞系,是EGFR及MET靶点单抗、双抗、联合治疗药物的体内药效及安全性评价的有力工具,期望助力全球药企研发进展。
部分数据展示
B-hEGFR/hMET小鼠
mRNA表达分析
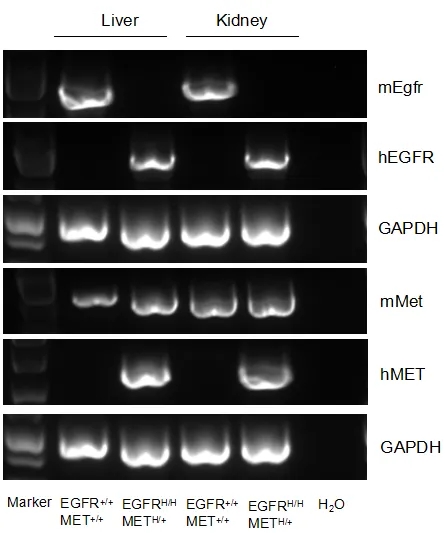
小鼠Egfr mRNA仅在野生型小鼠的肝脏和肾脏中检测到,小鼠Met mRNA在野生型小鼠和B-hEGFR/hMET小鼠的肝脏和肾脏中检测到。人EGFR mRNA及人MET mRNA仅在B-hEGFR/hMET小鼠中检测到,而在野生型小鼠中检测不到。
蛋白表达分析
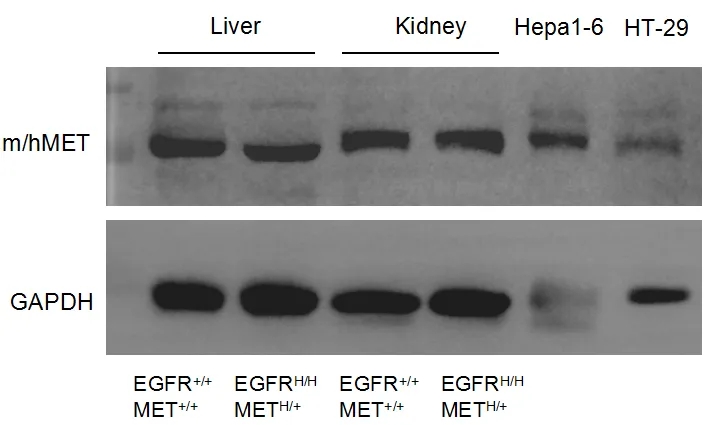
取野生型小鼠和B-hEGFR/hMET小鼠的肝脏和肾脏,用抗MET抗体进行western blot分析。由于抗体的交叉反应性,野生型小鼠和B-hEGFR/hMET小鼠均可检测到MET。
EGFR表达的IHC分析
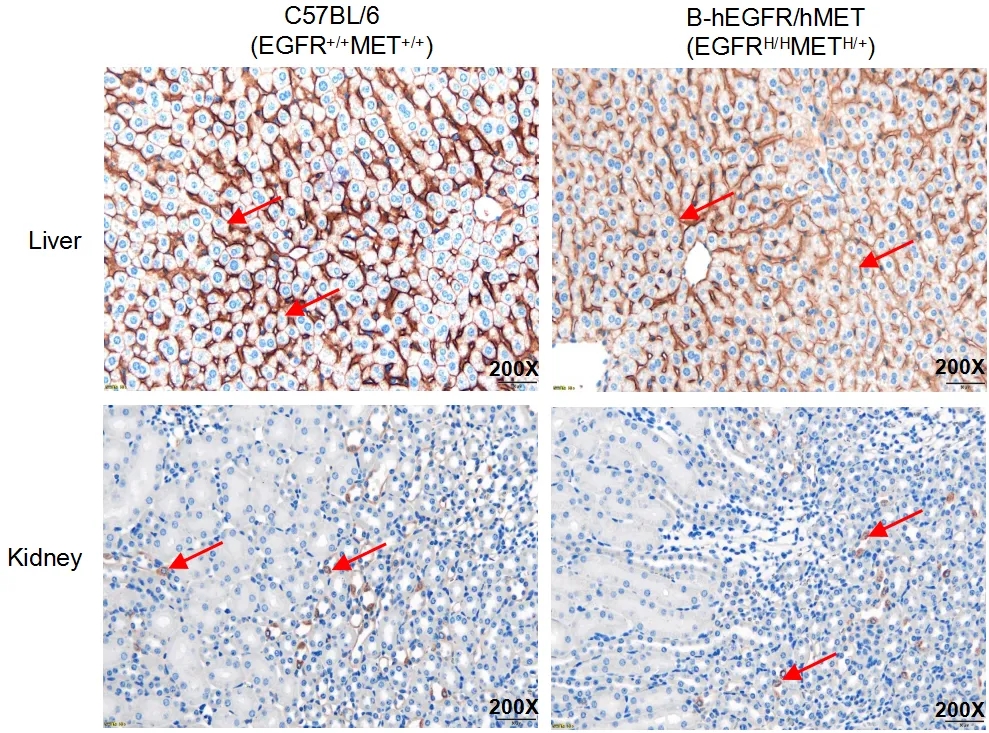
在野生鼠的肝脏和肾脏中检测到小鼠EGFR蛋白表达,在B-hEGFR/hMET小鼠中检测到人EGFR蛋白表达。
B-hEGFR小鼠及B-Tg(hEGFR) MC38细胞
EGFR表达的IHC分析
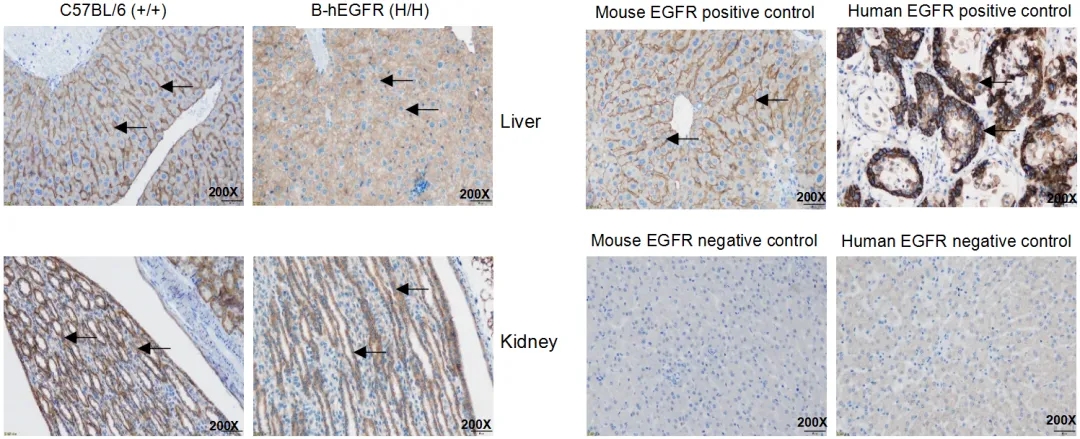
在野生鼠的肝脏和肾脏中检测到小鼠EGFR蛋白表达,在B-hEGFR小鼠中检测到人EGFR蛋白表达。箭头表示EGFR染色阳性的组织细胞(棕色)。
血常规/生化分析
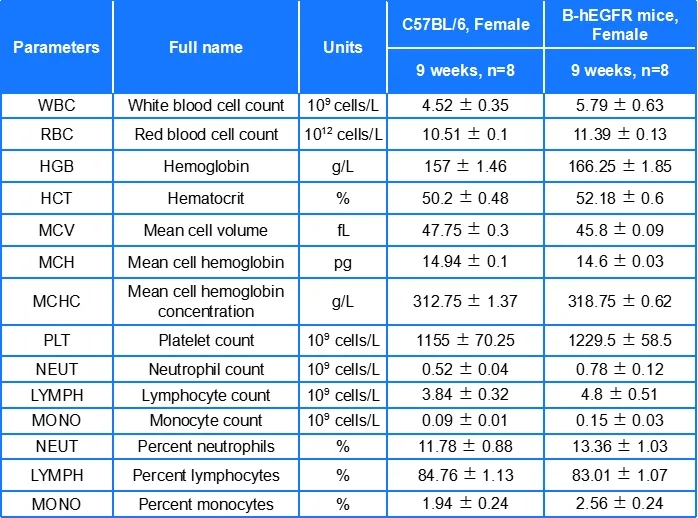
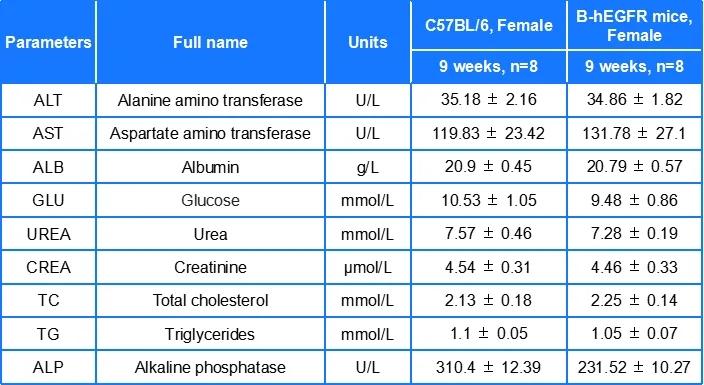
B-hEGFR小鼠各项血液参数与野生型小鼠相似,表明EGFR人源化没有影响小鼠血细胞的发育及生化指标。数值以平均值±SEM表示。
肿瘤生长曲线及体重变化
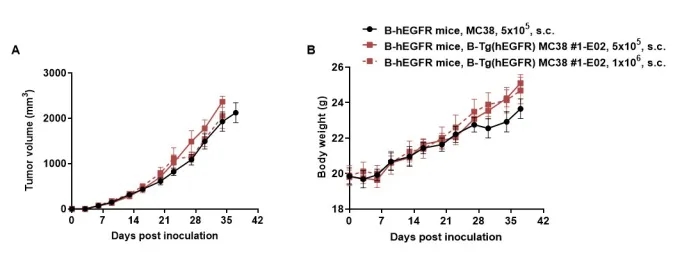
将B-Tg(hEGFR) MC38细胞(5×105,1×106)和野生型MC38细胞(5×105)皮下植入纯合子B-hEGFR小鼠(雌性,6-9周龄,n=6)。每周2次测量肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(Mean±SEM)。如图所示,B-Tg(hEGFR) MC38细胞在体内能够形成肿瘤,可用于疗效研究。体积以mm3表示,公式为:V=0.5×长径×短径2
肿瘤细胞蛋白表达分析
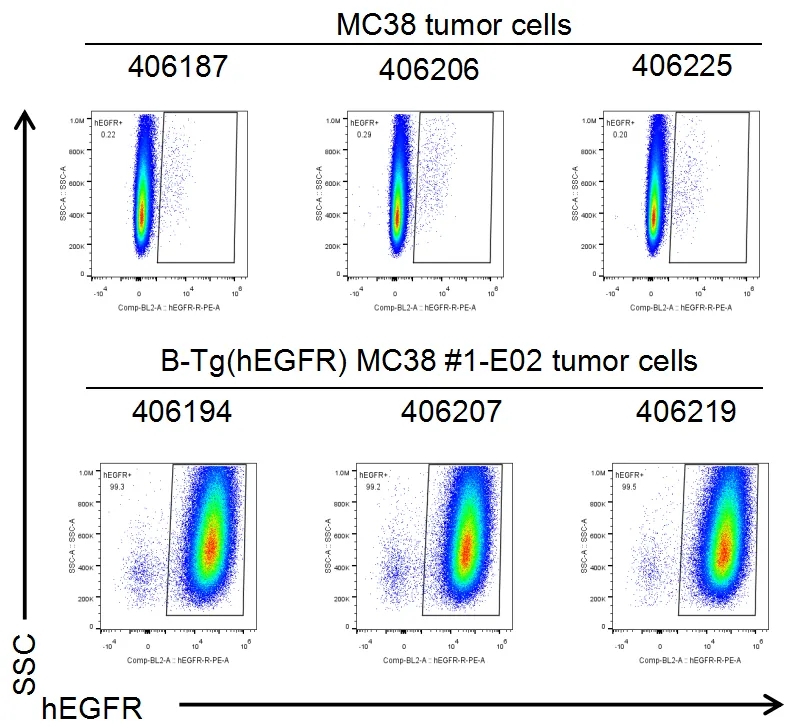
结试验时,取肿瘤组织检测人EGFR的表达情况,如图所示,人EGFR在肿瘤细胞表面高表达。
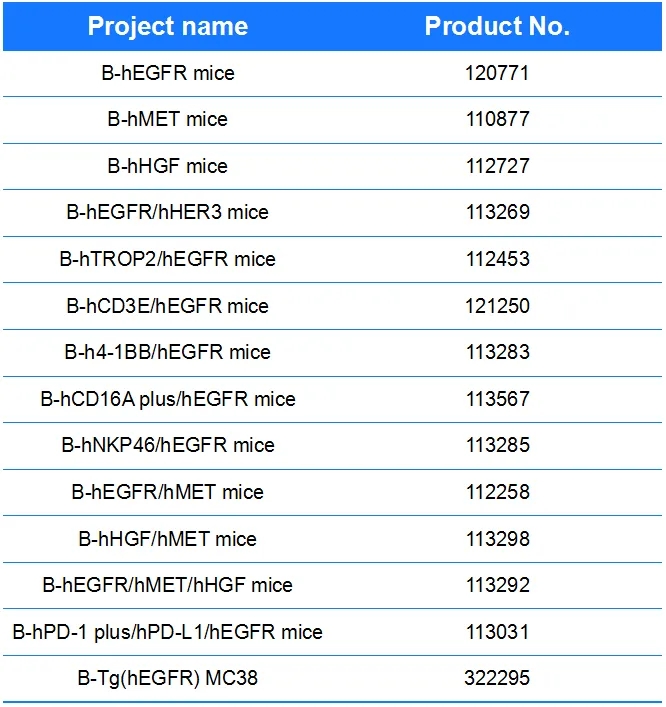
相关产品列表
参考资料:
[1] Kafkova LR, Mierzwicka JM, Chakraborty P, et al. NSCLC: from tumorigenesis, immune checkpoint misuse to current and future targeted therapy[J]. Front Immunol. 2024 Feb 7:15:1342086.
[2] Cai WQ, Zeng LS, Wang LF, et al. The Latest Battles Between EGFR Monoclonal Antibodies and Resistant Tumor Cells. Front Oncol. 2020;10:1249. Published 2020 Jul 24. doi:10.3389/fonc.2020.01249
[3] He J, Huang Z, Han L, et al. Mechanisms and management of 3rdgeneration EGFRTKI resistance in advanced nonsmall cell lung cancer[J]. International journal of oncology, 2021, 59(5): 1-20.
[4] Hong S, Gao F, Fu S, et al. Concomitant genetic alterations with response to treatment and epidermal growth factor receptor tyrosine kinase inhibitors in patients with EGFR‐ mutant advanced non‐small cell lung cancer. JAMA Oncol. 2018;4(5):739–42.
[5] Hang-Ping Yao et al, Progress and challenge in development of biotherapeutics targeting MET receptor for treatment of advanced cancer,Biochim Biophys Acta Rev Cancer . 2020 Dec;1874(2):188425.
[6] Wang Q, Yang S, Wang K and Sun SY: MET inhibitors for targeted therapy of EGFR TKI‑resistant lung cancer. J Hematol Oncol 12: 63, 2019.
[7] Drilon A, Cappuzzo F, Ou SI and Camidge DR: Targeting MET in lung cancer: Will expectations finally Be MET? J Thorac Oncol 12: 15‑26, 2017.
[8]Yun J, Lee SH, Kim SY, Jeong SY, Kim JH, et al. Antitumor Activity of Amivantamab (JNJ-61186372), an EGFR-MET Bispecific Antibody, in Diverse Models of EGFR Exon 20 Insertion-Driven NSCLC. Cancer Discov. 2020 Aug;10(8):1194-1209.





 010-56967680
010-56967680 info@bbctg.com.cn
info@bbctg.com.cn 苏公网安备:32068402320845号
网站建设:北京分形科技
苏公网安备:32068402320845号
网站建设:北京分形科技