
CD200分子被描述为一种自然产生的免疫调节剂,能够调节炎症和移植排斥反应,可以作为进展性恶性肿瘤的重要标志物。CD200诱导的调节在关节炎、过敏、感染和其他自身免疫性疾病中具有重要作用,可以促进感染、自身免疫性疾病和过敏等疾病的免疫调节。
除自免性疾病外,多项实验和临床研究已表明,CD200与其受体CD200R的相互作用导致多种免疫反应的衰减,降低对肿瘤生长的抵抗力,阻断CD200/CD200R信号通路是潜在的肿瘤免疫治疗手段[1]。
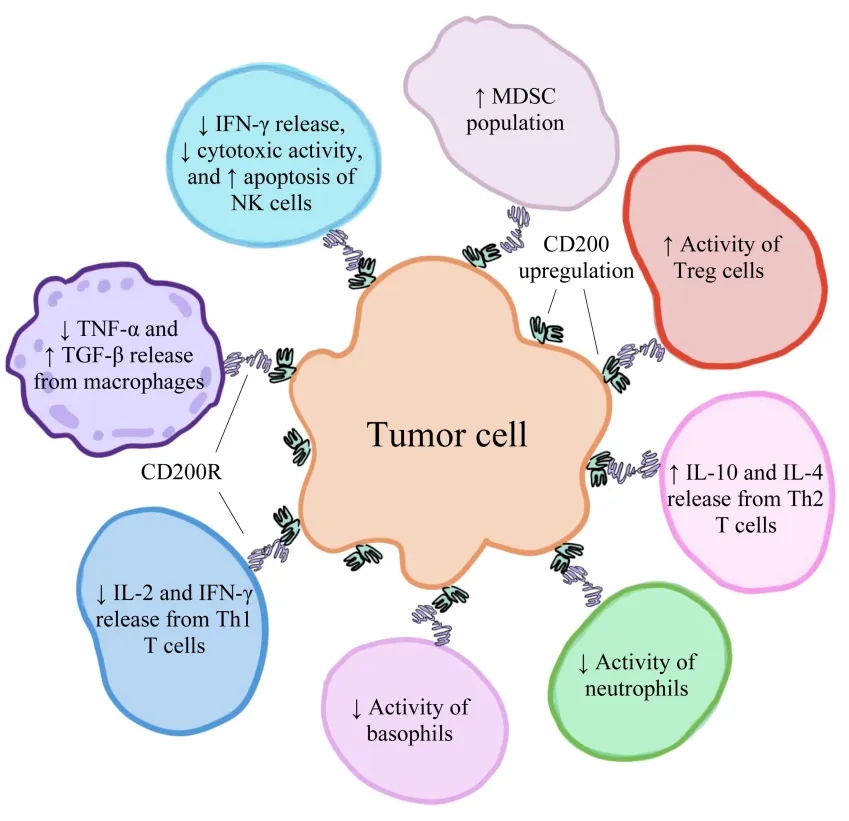
CD200/CD200R在各种人类癌细胞上对肿瘤微环境内的免疫细胞具有免疫抑制作用,创造有利于肿瘤发育、生长和扩散的条件[2]
CD200及CD200R靶点概述
CD200(也叫作 OX-2) 是 Ig 超家族(IgSF) 的一员, 具有与 B7 家族类似的结构。CD200 在包括胸腺细胞、 活化的 T 细胞、 B 细胞、 树突状细胞、 血管内皮细胞、 肺泡内皮细胞、 肾小球细胞、 平滑肌以及滋养层细胞等多种细胞上表达, 同时也是 CLL 或者 MM, CNS 相关肿瘤高表达的 Marker。CD200 的功能最初是通过缺乏 CD200 基因的小鼠来确定的, 在这些小鼠的自身免疫性脑脊髓炎(EAE) 、 胶原诱导的关节炎(CIA) 和自身免疫性脱发的实验模型中,小鼠较早的出现了疾病。小鼠脾脏巨噬细胞数量增加, 免疫刺激后, 这些巨噬细胞在体内表现出更活跃的表型。这些研究证实了 CD200 在维持自我耐受中的重要性, 表明 CD200 在控制自身免疫、 炎症和适应性免疫反应中起着重要的分子作用, 这意味着它是一个关键的免疫抑制分子。
CD200R 是 CD200 的受体, 同时也发现了多个与 CD200R 高度同源的家族分子(如 CD200RLa 等) , 这些分子胞外也是含有 Ig 结构域, 但是胞内没有明确的 motif 负责信号传递, 而是通过多个含有酪氨酸的 NPXY 序列通过磷酸化后启动下游信号途径。CD200R 的表达相对局限性较高, 主要是髓系细胞, 例如巨噬细胞, 单核细胞, DC 细胞等;CD200-CD200R 相互作用向髓系细胞发出抑制信号,在肿瘤TME 中的髓系细胞尤其是 MDSCs 等高表达CD200R 能够抑制抗肿瘤免疫,促进肿瘤生长。

CD200/CD200R的表达模式及胞内信号[3]
CD200/CD200R信号在部分疾病中的作用
异种移植
CD200可通过抑制CTL的发育,影响淋巴样细胞产生细胞因子的极化,细胞因子的极化增加2型细胞因子(IL-4、IL-10、TGFβ),减少1型细胞因子(IL-2、IFN-y、TNF-α),显著延长体内同种异体和异种移植模型的存活时间[4]。在CD200过表达的情况下,移植模型建立“耐受”状态后,在CD200不持续表达的情况下,Treg也可以持续表达维持移植模型存活。然而,炎症通常会导致Treg维持的稳定的移植模型发生排斥反应,CD200持续存在可以抑制这种验证导致的排斥反应[1]。
血液瘤
CD200作为一种表面抗原在骨髓/淋巴源性肿瘤细胞上的表达与许多人类血液系统恶性肿瘤的不良预后有关。据报道,经过高剂量化疗和自体造血干细胞移植,多发性骨髓瘤细胞(MMc)上过表达CD200的患者与MMc不过表达CD200的患者相比,其无事件生存期(EFS)更短[5]。在慢性淋巴细胞白血病(chronic lymphocytic leukemia, CLL)的研究中,阻断CLL中CD200的表达可降低CD4+CD25+Foxp3+Treg的数量,增强效应PBMCs产生的促炎细胞因子IFN-y和TNFa,并增强CD8+ CTL对肿瘤细胞的有效杀伤[6]。与MM和CLL的数据一致,CD200在毛细胞白血病(HCL)、ALL和AML病例中均过表达,并与该疾病的预后相关[7]。
实体瘤
CD200在实体肿瘤中广泛表达,如胰腺导管腺癌、肺癌、卵巢癌、肾细胞癌、中枢神经系统恶性肿瘤和头颈部鳞状癌(HNSCC),其与CD200R的相互作用促进肿瘤微环境中的免疫抑制。MDSC是未成熟的髓系细胞,在肿瘤细胞中高表达,抑制抗肿瘤免疫反应[8]。CD200/CD200R相互作用通过抑制巨噬细胞、诱导调节性T细胞、将细胞因子谱从Th1切换到Th2、抑制肿瘤特异性T细胞免疫和诱导MDSC(髓源性抑制细胞)扩增,从而促进免疫抑制[9]。
寄生虫/病毒感染
现在已知一些病毒表达一种CD200样蛋白,该蛋白被认为有助于在病毒感染后逃避宿主免疫[10]。来自疱疹病毒科的CD200样蛋白在结构上与宿主CD200最相似,预计它可以向宿主髓细胞传递调节信号并影响病毒感染的严重程度[11]。为了延长在宿主中生存时间,许多细菌和寄生虫病原体通过调节CD200或CD200R的表达,或通过表达CD200模拟物与宿主CD200R结合,从而减弱先天免疫。细菌和寄生虫生物通过CD200/CD200R相互作用减少先天细胞及其种群的促炎细胞因子释放,从而降低疾病的严重程度,延迟宿主病原体清除速度[12]。
自身免疫性疾病和炎症
胶原诱导关节炎(CIA)模型长期以来被用作类风湿关节炎(RA)的动物模型,促炎细胞因子,包括IFN-y、TNFα和IL-1ß,在CIA小鼠的关节炎关节中大量表达[13]。可溶性CD200-Fc融合蛋白或靶向CD200R单克隆抗体,可通过显著抑制对胶原的致敏作用阻止关节炎模型(CIA)的疾病发展,降低血清中TNFo和IFN-y,减少体外胶原致敏细胞释放相同的细胞因子[14]。
CD200+内皮细胞和神经元与CD200R+常驻巨噬细胞结合并阻止其经典活化,从而抑制Th1细胞因子反应,减轻自身免疫性葡萄膜视网膜炎模型中存在的炎症[15]。
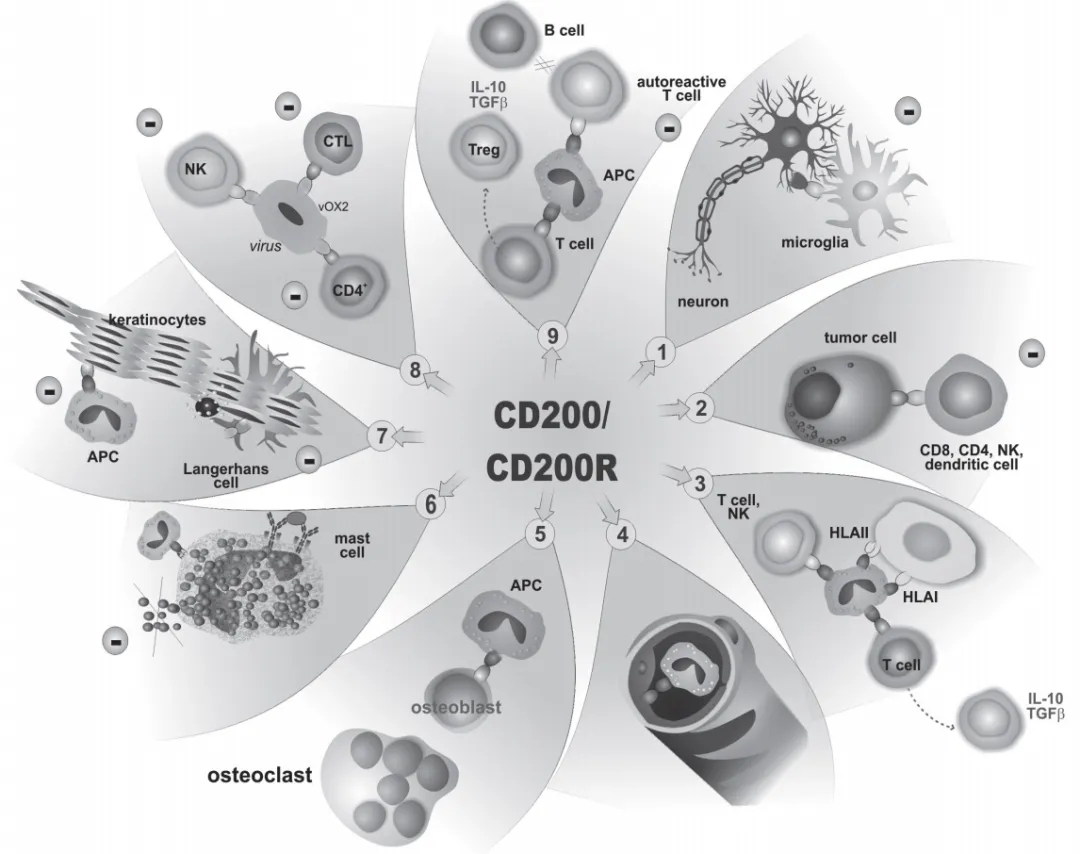
CD200/CD200R信号在不同疾病临床研究中的相互作用机制[16]
CD200/CD200R信号在自免疾病、炎症、肿瘤、神经系统疾病等多种疾病的免疫调节中起到重要作用,是潜在的免疫治疗靶点。百奥动物自主研发一系列CD200/ CD200R人源化小鼠及细胞系,助力相关免疫治疗药物研发。
CD200/CD200R人源化小鼠及细胞系
01、B-hCD200 mice
基本信息

蛋白表达分析
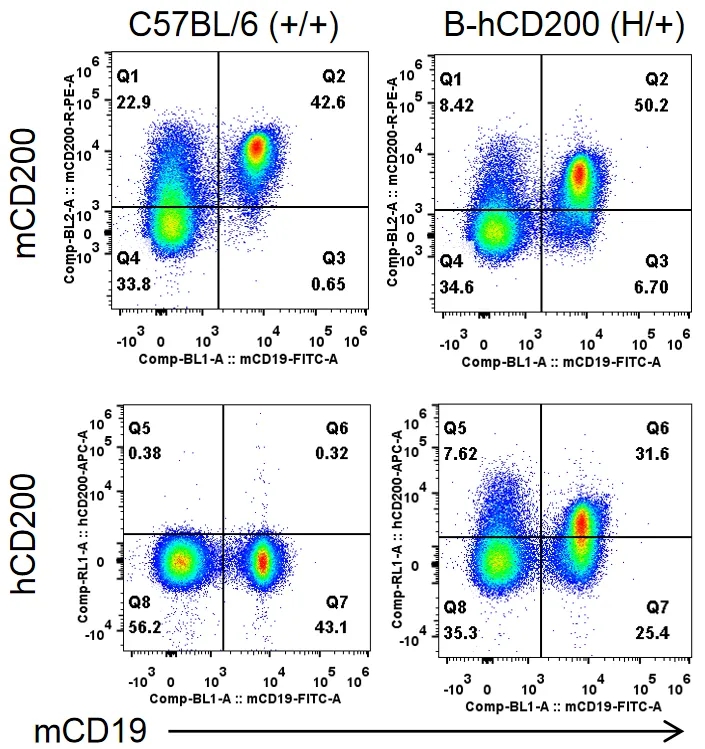
流式细胞术分析B-hCD200杂合小鼠中种属特异性CD200表达
收集野生型C57BL/6小鼠(+/+)和杂合B-hCD200小鼠(H/+)的脾细胞,用抗CD200抗体流式细胞术进行分析。小鼠CD200在C57BL/6小鼠和杂合B-hCD200小鼠中均可检测到。人CD200仅在杂合B-hCD200小鼠中检测到,而在C57BL/6小鼠中检测不到。
02、B-hCD200R mice
基本信息

蛋白表达分析
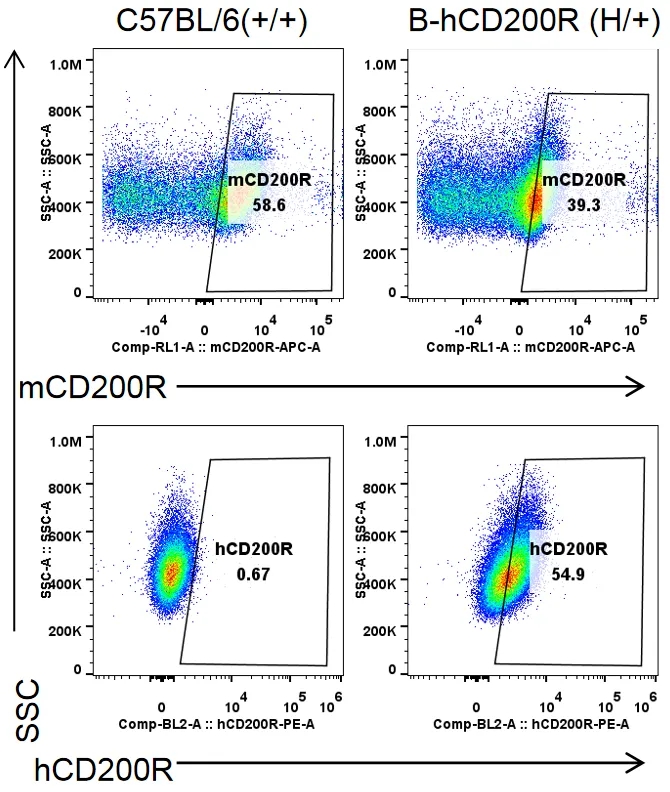
流式细胞术分析B-hCD200R杂合小鼠中种属特异性CD200R表达
收集野生型C57BL/6小鼠(+/+)和杂合B-hCD200R小鼠(H/+)腹膜巨噬细胞,用种属特异性抗CD200R抗体流式细胞术进行分析。小鼠CD200R在C57BL/6小鼠和杂合B-hCD200R小鼠中均可检测到。人CD200R仅在杂合B-hCD200R小鼠中检测到,而在C57BL/6小鼠中检测不到。
03、B-hCD200/hCD200R mice
基本信息

蛋白表达分析
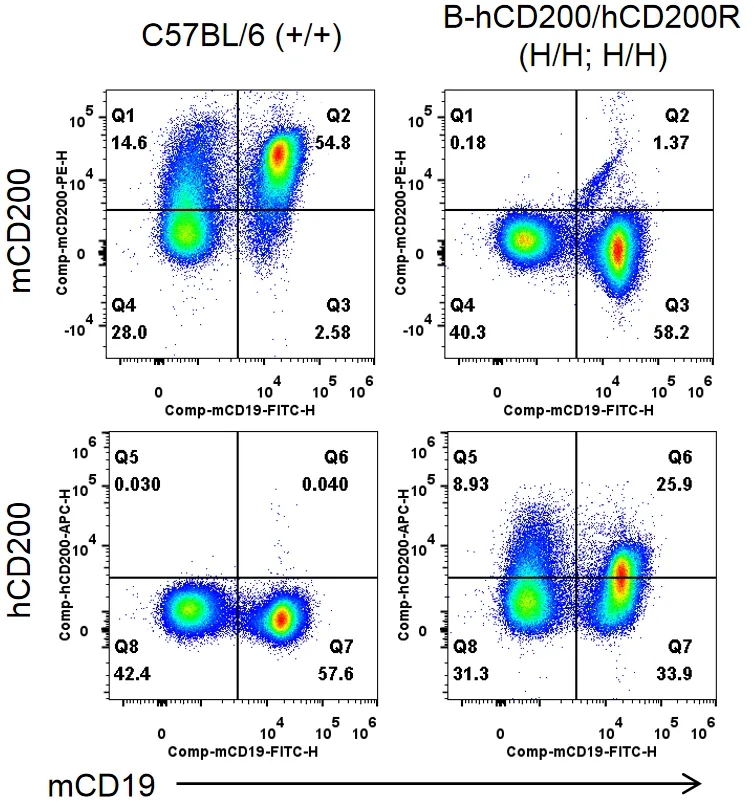
流式细胞术分析B-hCD200/hCD200R纯合小鼠种属特异性CD200表达
收集野生型C57BL/6小鼠(+/+)和纯合B-hCD200/hCD200R小鼠(H/H;H/H)的脾细胞,用种属特异性抗CD200抗体流式细胞术进行分析。小鼠CD200在C57BL/6小鼠中检测到。人CD200仅在纯合B-hCD200/hCD200R小鼠中检测到,而在C57BL/6小鼠中检测不到。
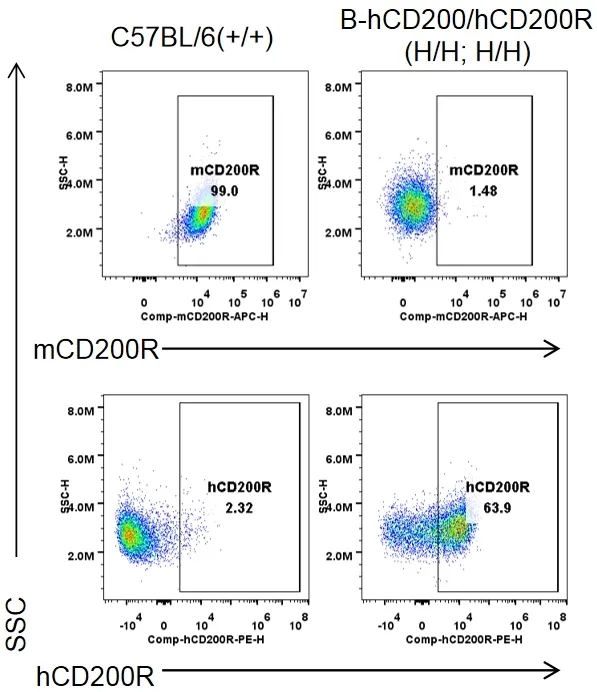
流式细胞术分析B-hCD200/hCD200R纯合小鼠种属特异性CD200R表达
收集野生型C57BL/6小鼠(+/+)和纯合B-hCD200/hCD200R小鼠(H/H;H/H)腹膜巨噬细胞,用种属特异性抗CD200R抗体流式细胞术进行分析。小鼠CD200R在C57BL/6小鼠中检测到。人CD200R仅在纯合B-hCD200/hCD200R小鼠中检测到,而在C57BL/6小鼠中检测不到。
免疫分型
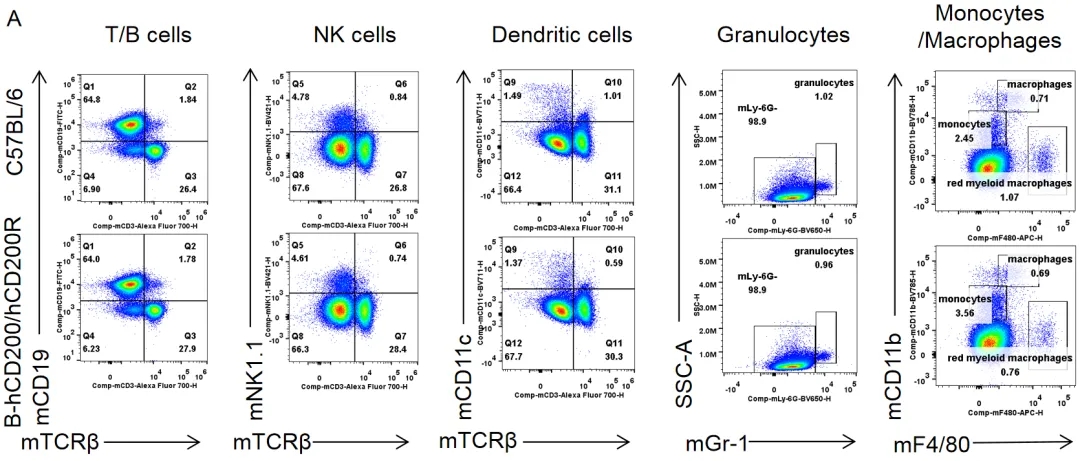
脾脏白细胞亚群的流式细胞术分析
从雌性C57BL/6和纯合B-hCD200/hCD200R小鼠(n= 3,8周龄)中分离脾细胞。用流式细胞术分析脾细胞以评估白细胞亚群。纯合B-hCD200/hCD200R小鼠的T细胞、B细胞、NK细胞、树突状细胞、粒细胞、单核细胞和巨噬细胞的百分比与C57BL/6小鼠相似,表明人源化CD200/CD200R并不会改变脾脏中这些白细胞亚群的整体发育、分化或分布。数值以平均值±SEM表示。
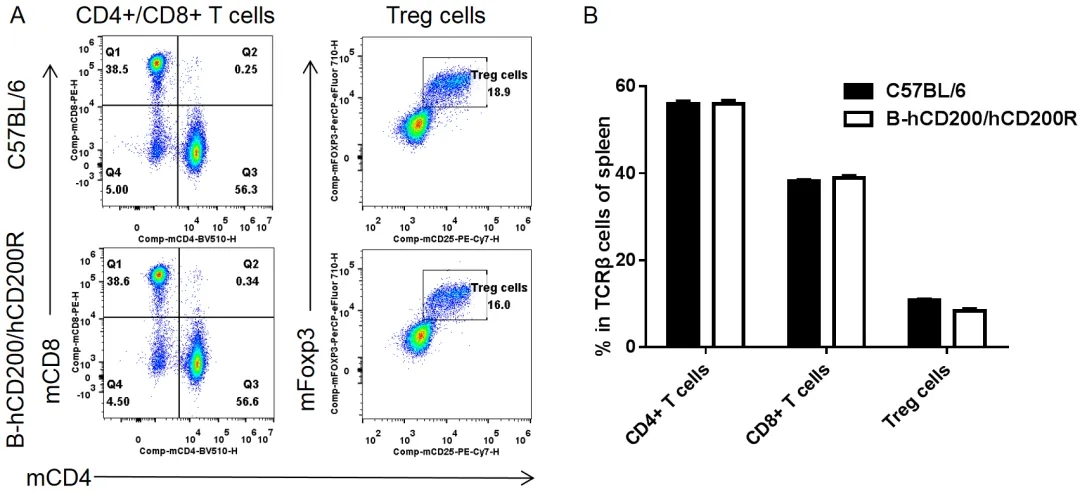
脾脏T细胞亚群的流式细胞术分析
从雌性C57BL/6和纯合B-hCD200/hCD200R小鼠(n= 3,8周龄)中分离脾细胞。用流式细胞术分析脾细胞以评估T细胞亚群。纯合B-hCD200/hCD200R小鼠中CD8+ T细胞、CD4+ T细胞和Treg细胞的百分比与C57BL/6小鼠相似,这表明人源化CD200/CD200R并不会改变脾脏中这些T细胞亚群的整体发育、分化或分布。数值以平均值±SEM表示。
实验结果表明人源化CD200/CD200R同样也不影响淋巴结中白细胞亚群和T细胞亚群总体的发育、分化和分布。(数据未展示)
体内药效
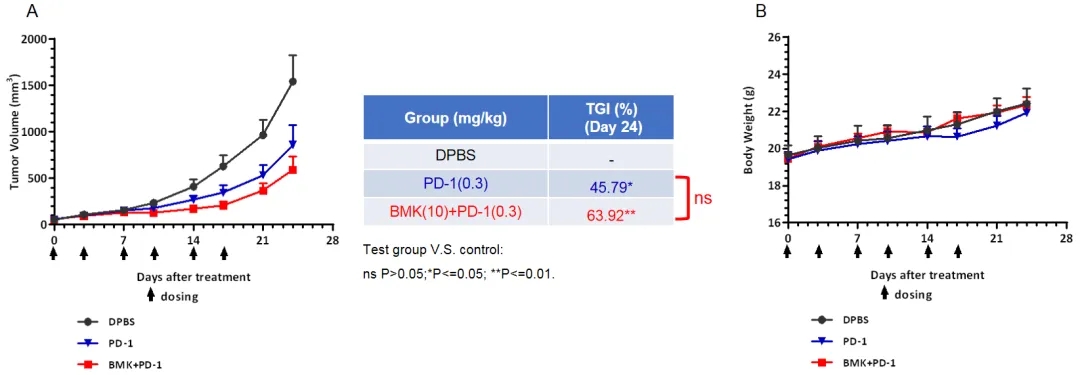
抗人CD200 (hCD200)抗体与抗小鼠PD-1 (mPD-1)抗体联合使用时的体内抗肿瘤活性
将B-hCD200 MC38细胞(5x105)皮下植入纯合B-hCD200/hCD200R小鼠(雌性,6-8周龄,n=6)体内。当肿瘤体积达到约100 mm3时,将小鼠分组进行治疗。结果显示,抗hCD200抗体联合抗mPD-1抗体可有效控制B-hCD200/hCD200R小鼠的肿瘤生长,说明B-hCD200/hCD200R小鼠为体内评价抗人CD200抗体提供了强有力的临床前模型。数值以平均值±SEM表示。采用双因素方差分析(ANOVA)检验。ns P>0.05;*P<=0.05; **P<=0.01。
注:本实验由客户使用B-hCD200/hCD200R小鼠进行。所有其他材料均由客户提供。
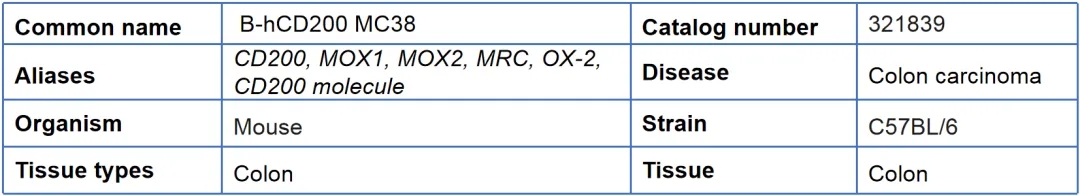
04、B-hCD200 MC38
基本信息
细胞系成瘤实验
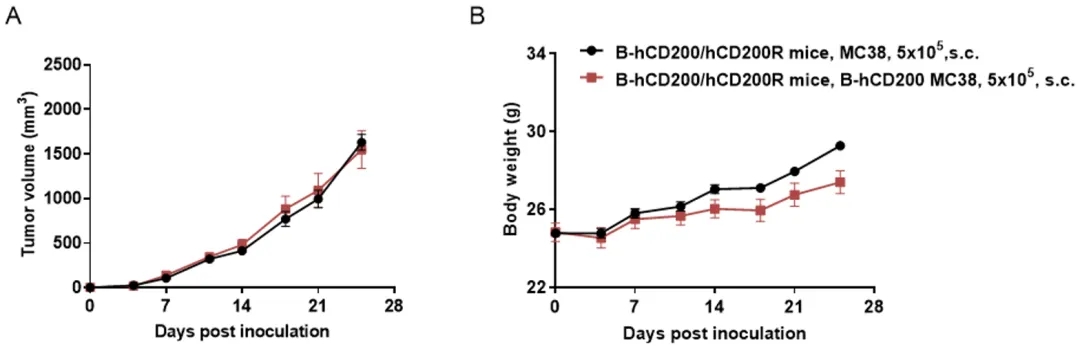
将B-hCD200 MC38细胞(5x105)和野生型MC38细胞(5x105)皮下植入B-hCD200/hCD200R小鼠体内(雄性,7周龄,n=6)。每周2次测量肿瘤体积和体重。(A)平均肿瘤体积±SEM。(B)体重(平均值±SEM)。体积以mm3表示,公式为:V=0.5×长径×短径2。如A图所示,B-hCD200 MC38细胞在体内能够形成肿瘤,可用于临床前药效。
肿瘤细胞蛋白表达分析
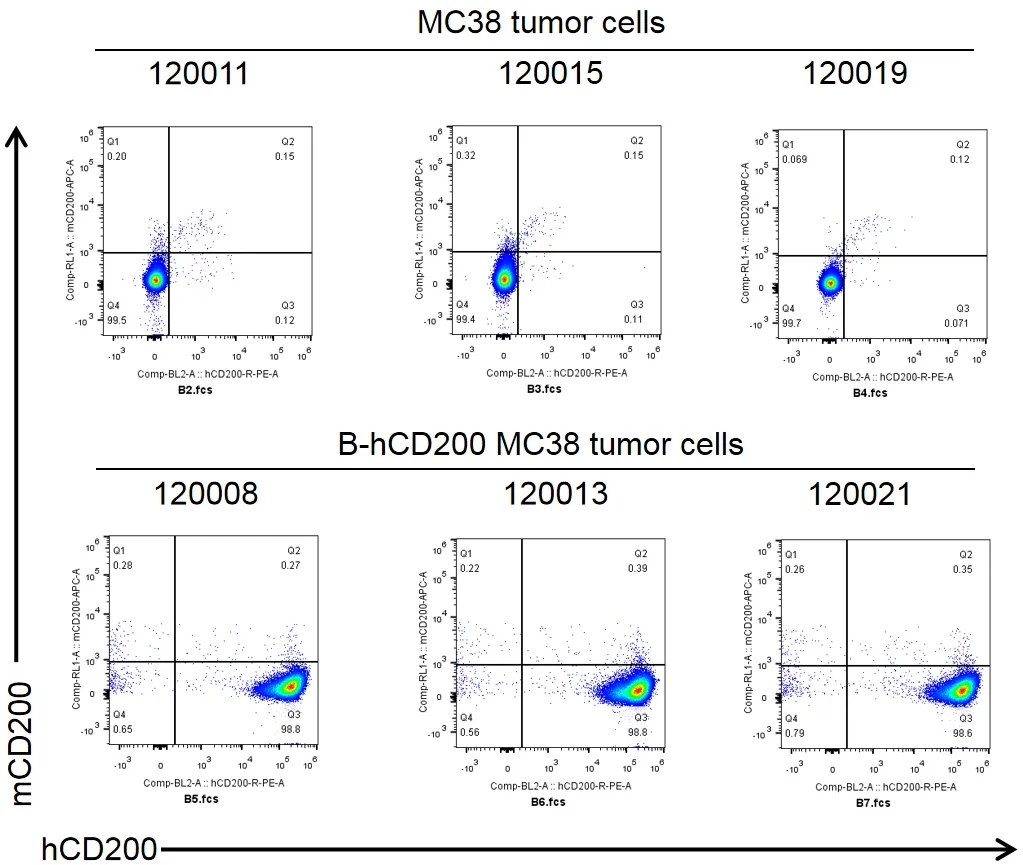
将B-hCD200 MC38细胞皮下移植到B-hCD200/hCD200R小鼠体内(n=6)。实验结束时,收集肿瘤细胞,用流式细胞术检测人CD200的表达。如图所示,人CD200在肿瘤细胞表面高度表达。因此,B-hCD200 MC38细胞可用于体内疗效研究。
注意,B-hCD200 MC38细胞仅在B-hCD200/hCD200R小鼠中成功形成肿瘤,而在野生型小鼠中不成瘤。
参考资料
[1]Kotwica-Mojzych, K.;Jodłowska-J˛edrych, B.; Mojzych, M.CD200:CD200R Interactions and Their Importance in Immunoregulation. Int. J. Mol. Sci.2021, 22, 1602.
[2]Choe D, Choi D. Cancel cancer: The immunotherapeutic potential of CD200/CD200R blockade. Front Oncol. 2023 Jan 23;13
[3]Tomasz P Rygiel,Linde Meyaard. CD200R signaling in tumor tolerance and inflammation:A tricky balance.Current Opinion in Immunology 2012, 24:233–238.
[4]Gorczynski, R.M. CD200 and its receptors as targets for immunoregulation. Curr. Opin. Investig. Drugs. Rev. 2005, 65, 483–488.
[5]Moreaux, J.; Hose, D.; Reme, T.; Jourdan, E.; Hundemer, M.; Legouffe, E.; Moine, P.; Bourin, P.; Moos, M.; Corre, J.; et al. CD200 is a new prognostic factor in multiple myeloma. Blood 2006, 108, 4194–4197.
[6]Wong, K.K.; Khatri, I.; Shaha, S.; Spaner, D.E.; Gorczynski, R.M. The role of CD200 in immunity to B cell lymphoma. J. Leukoc.Biol. 2010, 88, 361–372.
[7]Aref, S.; Azmy, E.; El-Bakry, K.; Ibrahim, L.; Mabed, M. Prognostic impact of CD200 and CD56 expression in adult acute lymphoblastic leukemia patients. Hematology 2018, 23, 263–270.
[8]Mundy-Bosse, B.L.; Lesinski, G.B.; Jaime-Ramirez, A.C.; Benninger, K.; Khan, M.; Kuppusamy, P.; Guenterberg, K.; Kondadasula,S.V.; Chaudhury, A.R.; La Perle, K.M.; et al. Myeloid-derived suppressor cell inhibition of the IFN response in tumorbearing mice.Cancer Res. 2011, 71, 5101–5110.
[9]Choueiry, F.; Torok, M.; Shakya, R.; Agrawal, K.; Deems, A.; Benner, B.; Hinton, A.; Shaffer, J.; Blaser, B.W.; Noonan, A.M.;et al. CD200 promotes immunosuppression in the pancreatic tumor microenvironment. J. Immunother Cancer 2020, 8, e000189.
[10]Chen, Z.; Ma, X.; Zhang, J.; Hu, J.; Gorczynski, R.M. Alternative splicing of CD200 is regulated by an exonic splicing enhancer and SF2/ASF. Nucleic Acids Res. 2010, 38, 6684–6696.
[11]Foster-Cuevas, M.; Wright, G.J.; Puklavec, M.J.; Brown, M.H.; Barclay, A.N. Human herpesvirus 8 K14 protein mimics CD200 in down-regulating macrophage activation through CD200 receptor. J. Virol. 2004, 78, 7667–7676.
[12]Vaine, C.A.; Soberman, R.J. Signaling Pathway: Immune Regulation and Host–Pathogen Interactions. Adv. Immunol. 2014,21, 191–211.
[13]Luross, J.A.; Williams, N.A. The genetic and immunopathological processes underlying collagen-induced arthritis. Immunology 2001, 103, 407–416.
[14]Egle Sˇimelyte, Gabriel Criado,1 David Essex,Robert A. Uger, Marc Feldmann,and Richard O. Williams.CD200-Fc, a Novel Antiarthritic Biologic Agent That Targets Proinflammatory Cytokine Expression in the Joints of Mice With Collagen-Induced Arthritis.ARTHRITIS & RHEUMATISM Vol. 58, No. 4, April 2008, pp 1038–1043
[15]Dick, A.D.; Carter, D.; Robertson, M.; Broderick, C.; Hughes, E.; Forrester, J.V.; Liversidge, J. Control of myeloid activity during retinal inflammation. J. Leukoc. Biol. 2003, 74, 161–166.
[16]Drahomíra Holmannová, Martina Koláčková, Kateřina Kondělková, Pavel Kuneš1, Jan Krejsek, Ctirad Andrýs.CD200/CD200R PAIRED POTENT INHIBITORY MOLECULES REGULATING IMMUNE AND INFLAMMATORY RESPONSES; PART II: CD200/CD200R POTENTIAL CLINICAL APPLICATIONS.ACTA MEDICA (Hradec Králové) 2012; 55: 59-65.





 010-56967680
010-56967680 info@bbctg.com.cn
info@bbctg.com.cn 苏公网安备:32068402320845号
网站建设:北京分形科技
苏公网安备:32068402320845号
网站建设:北京分形科技